# Crear la tabla
chi_tabla <- data.frame(
gl = 1:5,
p_095 = c(0.004, 0.10, 0.35, 0.71, 1.14),
p_090 = c(0.02, 0.21, 0.58, 1.06, 1.61),
p_080 = c(0.06, 0.45, 1.01, 1.65, 2.34),
p_070 = c(0.15, 0.71, 1.42, 2.20, 3.00),
p_050 = c(0.46, 1.39, 2.37, 3.36, 4.35),
p_030 = c(1.07, 2.41, 3.66, 4.88, 6.06),
p_020 = c(1.64, 3.22, 4.64, 5.99, 7.29),
p_010 = c(2.71, 4.60, 6.25, 7.78, 9.24),
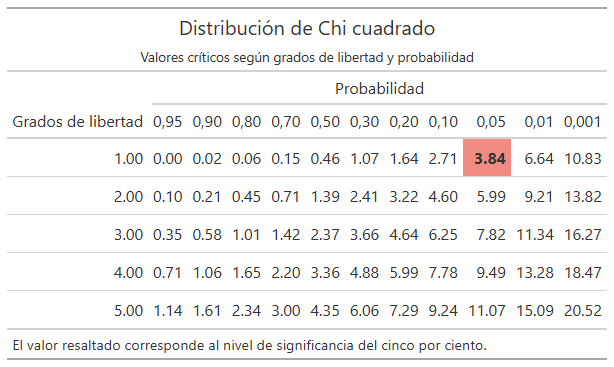
p_005 = c(3.84, 5.99, 7.82, 9.49, 11.07),
p_001 = c(6.64, 9.21, 11.34, 13.28, 15.09),
p_0001 = c(10.83, 13.82, 16.27, 18.47, 20.52)
)
# Tabla gt didáctica
chi_tabla %>%
gt() %>%
cols_label(
gl = "Grados de libertad",
p_095 = "0,95",
p_090 = "0,90",
p_080 = "0,80",
p_070 = "0,70",
p_050 = "0,50",
p_030 = "0,30",
p_020 = "0,20",
p_010 = "0,10",
p_005 = "0,05",
p_001 = "0,01",
p_0001 = "0,001"
) %>%
tab_header(
title = "Distribución de Chi cuadrado",
subtitle = "Valores críticos según grados de libertad y probabilidad"
) %>%
fmt_number(decimals = 2) %>%
tab_style(
style = list(
cell_fill(color = "#f28b82"),
cell_text(weight = "bold")
),
locations = cells_body(
rows = gl == 1,
columns = p_005
)
) %>%
tab_spanner(
label = "Probabilidad",
columns = p_095:p_0001
) %>%
tab_source_note(
source_note = "El valor resaltado corresponde al nivel de significancia del cinco por ciento."
)